
4D Cell-ECM interactions in multicellular systems
私たちの研究室では、細胞外マトリックス(ECM)を細胞の単なる静的な足場として捉えるのではなく、細胞集団を統率する細胞外の動的な情報マトリックスとして位置づけ、それが組織の形態形成および再生を制御する機構を明らかにすることを目指しています。具体的には、哺乳類の皮膚と初期胚/初期胚モデルを主な研究対象とし、ECMと細胞集団の空間マッピング、定量的な4D(3D +時間)イメージング、時間分解能をもつ単一細胞トランスクリプトーム解析、そして多様な摂動実験を組み合わせ、ECMの分子組成・構造・力学特性の時空間変化が組織をどのように形作るのかを解明しています。私たちの長期的な目標は、細胞とECMとの相互作用を設計・操作することで、形態形成やパターニングを細胞外から自在に制御することです。

1. ECM spatiotemporal code
Spatiotemporal information encoded in the ECM
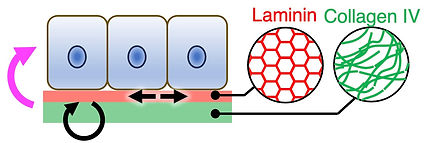
組織幹細胞はどのようなECM環境の中で維持され、機能しているのでしょうか?この問いに取り組むため、私たちは毛包をモデルとしてECMタンパク質の大規模空間マッピングを行い、幹細胞やその周囲のニッチ細胞が接着する基底膜の分子組成を網羅的に解析してきました。その結果、基底膜を構成するECM分子が時空間的に高度なパターンを形成していることを見出しました。さらに、これらのECM分子が異なる上皮幹細胞/前駆細胞とニッチ細胞との相互作用を選択的に仲介することで、組織の発生や再生を制御していることを明らかにしてきました (Fujiwara et al., Cell 2011; Cheng et al., eLife 2018; Tsutsui et al., Nat Commun 2021; Liu et al., J Invest Dermatol 2025; Yokota et al., Cell Rep 2025など)。現在は、ECMタンパク質の空間マッピングと空間トランスクリプトミクスを統合することで、ECMの分子パターン(ECMコード)と細胞状態との関係を体系的に解明する研究へと発展しています。また、得られた知見をもとに、試験管内でECMニッチを再構成・設計し、組織幹細胞やニッチ細胞を制御する研究にも取り組んでいます (Machida et al., Matrix Biol 2026)。

2. ECM dynamics
Multi-scale ECM dynamics in 4D
ECMは長らく、細胞を支える静的な足場として捉えられてきました。しかし近年、ECMは絶えず再編成される動的なポリマーネットワークであり、その構造や力学特性を通じて組織形成や再生に重要な役割を果たすことが徐々に明らかになってきています。一方で、ECMの分子動態がどのように制御され、それがどのように組織レベルの形態形成へと結びつくのかについては、未だ十分に理解されていません。この問題に取り組むため、私たちは、生体内の基底膜を長時間・定量的に4次元観察できる蛍光基底膜レポーターノックインマウスを開発しました (Wuergezhen et al., J Cell Biol 2025)。その解析から、基底膜は一様で静的なシート構造ではなく、コラーゲンIVの分子ターンオーバーが時空間的に制御された動的なポリマーネットワークであることを見出しました。さらに、この分子動態が基底膜のマクロな拡張や上皮組織の異方的成長と密接に連動していることも明らかにしました。現在は、ライブイメージングによって得られる定量データ、組織内の力学計測、数理モデリングを統合し、分子ターンオーバー、組織スケールでの基底膜の拡張、そして力学特性との関係をポリマーダイナミクスの観点から記述・予測する理論基盤の構築を進めています。最終的には、基底膜を自己組織化するアダプティブ・ポリマーネットワークとして理解し、その動態が組織力学をどのように調節するのか、さらに細胞との相互作用を通じてどのように新たな組織構造や形態を創発するのか、その原理の解明を目指しています。

3. Development & regeneration
Emergence of tissue architecture through cell–ECM interactions
皮膚は、生物と環境とのインターフェースとして機能します。多様な生態環境へ適応するために、皮膚はその構造と機能を多様化させてきました。毛包、汗腺、乳腺、羽毛、ウロコなどの皮膚付属器は、その多様性を生み出す主要な要素です。興味深いことに、これらの器官は、発生初期にはプラコードと呼ばれる類似した円盤状の上皮構造から形成されますが、その後、それぞれ異なる3次元形態を獲得します。さらに、生涯にわたる維持や再生を可能にするため、発生過程で組織幹細胞を形成します。しかし、器官ごとの形態的多様性と再性能の獲得が、どのような共通原理によって実現されるのかは十分に解明されていません。
この問題を理解するためには、器官がどのような形態形成過程を経て構築されるのかを明らかにするだけでなく、その過程で細胞状態や相互作用がどのように変化し、将来の機能や運命へと結びつくのかを時空間的に解析する必要があります。さらに、その理解を異なる器官間で比較できる形で記述することも重要です。
そこで私たちは、マウス毛包の発生動態を連続的に捉えるために、1細胞解像度の4Dイメージングと時系列シングルセルトランスクリプトーム解析を組み合わせたマルチスケール・マルチモーダル時空間解析を行いました (Morita et al., Nature 2021)。その結果、毛包上皮は、発生初期のプラコードに形成される2次元の同心円状プレパターンを出発点として、円筒状機能ドメインを持つ3次元形態へと段階的に構築されることを見出しました。さらにこのプレパターンの最外層領域が将来の毛包上皮幹細胞の起源であることも明らかにしました。私たちは、この「同心円から円筒へ」の協調的な発生過程を「テレスコープ型形態形成」と呼んでいます。このモデルでは、細胞集団とその周囲の細胞外環境が段階的に区画化されながら時空間発展することで、器官の立体構造形成と幹細胞ニッチの構築が協調的に進行すると考えています。現在は、この概念を乳腺や羽毛など他の器官へ適用し、器官間および種間比較を通じて検証しています。将来的には、発生・再生・進化に共通する皮膚付属器間の形態形成の普遍原理と、その改変によって多様な形態や再生能力が創発される仕組みの解明を目指しています。


4. Technology & tool development
Quantitative 4D mapping, imaging and manipulation across scales
上記の研究は、私たちが独自に開発してきた、細胞集団やECMを単一細胞解像度で4次元的に可視化・定量化する技術基盤によって支えられています。私たちは、細胞の動きや運命、ECMの構造や動態を生きた組織の中で長期間観察し、それらを統合的に解析するためのさまざまな技術を開発してきました。現在も、新しいイメージング技術、人工タンパク質、遺伝子操作技術などの開発を進めており、将来的には細胞とECMとの相互作用を自在に設計・操作することを目指しています。
主な技術・ツール
-
ECMアトラス構築技術:基底膜を構成するECM分子の種類や配置を組織内で網羅的に可視化し、空間マップとして記述する技術 (Tsutsui et al., Nat Commun 2021)
-
単一細胞解像度の4Dイメージングと時系列シングルセルトランスクリプトーム統合解析:発生過程における細胞の移動、分裂、運命決定を長期間追跡し、細胞の状態変化と結びつけて解析する技術 (Morita et al., Nature 2021)
-
生体内ECMのライブイメージング:生体組織内の基底膜をリアルタイムかつ定量的に可視化し、その動態を分子から組織のスケールで4次元解析できるノックインマウス (Wuergezhen et al., J Cell Biol 2025)
-
単一細胞トランスクリプトームによる再生ダイナミクス解析:周期的な組織再生の進行過程を単一細胞レベルで再構成し、再生過程の全体像を時系列で解析する技術(応用例:疑似毛周期の構築)(Yokota et al., Cell Rep 2025)。
Our goal
私たちの長期的なゴールは、細胞外微小環境がどのような原理で機能しているのかを解明し、その理解に基づいて微小環境を設計・操作することで、細胞の外側から生命現象を自在に制御することです。